So, Europe has a brand-new species of hitherto-undiscovered mammal, the
So, Europe has a brand-new species of hitherto-undiscovered mammal, the Cypriot mouse
Mus cypriacus.
That’s great, but what has interested me in particular is the claim made in many articles that the Cypriot mouse is ‘the first new European mammal to be discovered in more than 100 years’ (go, for example, here or here). Sad to say, this quote wasn’t invented by journalists, but apparently comes right from the mammalogists who described the species. One internet article on the discovery states that it ‘overturns the widely held belief that every living species of mammal had been identified in Europe’, and goes on to state that ‘it was generally assumed that the European biodiversity had been entirely picked over by the natural history pioneers of the 19th century’. Well, ok, something can be a ‘widely held belief’ and still be pretty much untrue, but while one might expect that Europe is a well known place where few new species are found nowadays, these statements – like most media statements pertaining to the rarity of recently discovered species – are wildly inaccurate. Sure, there aren’t as many new mammals coming out of 21st century Europe as there are frogs coming out of Sri Lanka or whatever, but the fact remains that Europe – the most well-explored and intensively studied continent of them all – most certainly has produced new mammal species within the last 100 years, including within recent decades. What’s more, it hasn’t produced one or two new species, but 32 of them! Sorry Mus cypriacus, but you ain’t that special.
You will know from previous blog posts that during the last few decades a large number of tropical rodents have been named and described (see New, obscure, and nearly extinct rodents of South America and Giant furry pets of the Incas). And so it is with Europe, and among rodents we start with mice. Remember here that we’re only interested in those species that have been named over the past 100 years.
Five European mice have been named within the last 100 years, four of which are obscure, and one of which is well known and well studied. Firstly, we have the Cretan spiny mouse Acomys minous Bate, 1906, a cold-adapted island endemic (Bate’s publication is sometimes given as 1905, in which case this isn’t a ‘100 year’ mammal). The second species, the Western house mouse Mus domesticus Schwartz & Schwartz, 1943, is anything but poorly known, and though not recognized as distinct from M. musculus Linnaeus, 1758 until 1943, it can hardly be regarded as a recently discovered species. As the common name suggests, the Western house mouse is the house mouse species of western Europe (as well as northern Africa). It is replaced in Scandinavia and eastern Europe by M. musculus, and around the Mediterranean coast it is sympatric with the Algerian mouse M. spretus.
The third ‘100 year’ European mouse species was first described from Allgäu in Germany: it’s the Alpine wood mouse Apodemus alpicola Heinrich, 1952, now known to occur in the Alps of Switzerland, Liechenstein, Austria and Italy as well as those of Germany. Though first named as a new species, it later became regarded as a high-altitude subspecies of the Yellow-necked mouse A. flavicollis. A 1989 study demonstrated that it should be recognised as a distinct species again. Also belonging to the genus Apodemus is the Mount Hermon field mouse A. iconicus Heptner, 1948. This species has a complex nomenclatural history that I won’t cover in full here, but of special interest is that a new species named from Israel in 1989 (A. hermonensis Filippucci et al., 1989) is now thought to be a junior synonym of A. iconicus: there are also a few older names (Mus sylvaticus var. tauricus Pallas, 1811, M. s. tauricus Barrett-Hamilton, 1900 and M. s. witherbyi Thomas, 1902) that some mammalogists regard as senior synonyms of A. iconicus, and if this is correct then A. iconicus is not a ‘100 year’ mammal. Though best associated with Israel and Turkey, A. iconicus has recently been added to the definitive European list as it’s now known to occur on Rhodes and Bozcaada (Kryštufek & Mozetič Francky 2005). Finally among mice, there is the recently discovered and poorly known Balkan short-tailed mouse Mus macedonicus Petrov & Ruzic, 1983.
Exactly as obscure as some of these mice are various ‘100 year’ vole species. One of them is comparatively well known however, and indeed is the best known recently discovered European mammal: the Bavarian pine vole Microtus bavaricus Konig, 1962 of the Bavarian and Italian Alps. Ironically, the reason the species is ‘best known’ is because it was thought to have become extinct: there was an absence of sightings after its discovery, and in 1980 a hospital was constructed on the location where it formerly occurred. However, the species was rediscovered by Friederike Spitzenberger in 2004 at a location in Austria.
 Four other Microtus voles have been named within the last 100 years. Cabrera’s vole Microtus cabrerae Thomas, 1906 is a poorly known, endangered Spanish species. Far better studied is the Sibling vole Microtus rossiaemeridionalis Ognev, 1924, a species that occurs from Finland southward to Greece, and also occurs in eastern Asia (see adjacent image). Between 30 and 70 years ago it was accidentally introduced to Svalbard, and in some years large numbers of the species occur there. Though originally named in the 1920s, the name M. rossiaemeridionalis was forgotten about in the following decades. The discovery in the late 1960s that a population originally assumed to be part of the Common vole M. arvalis actually merited distinction then led to the naming of the new species M. subarvalis Meyer et al. 1972, and it was this population that then proved to be the same thing as M. rossiaemeridionalis.
Four other Microtus voles have been named within the last 100 years. Cabrera’s vole Microtus cabrerae Thomas, 1906 is a poorly known, endangered Spanish species. Far better studied is the Sibling vole Microtus rossiaemeridionalis Ognev, 1924, a species that occurs from Finland southward to Greece, and also occurs in eastern Asia (see adjacent image). Between 30 and 70 years ago it was accidentally introduced to Svalbard, and in some years large numbers of the species occur there. Though originally named in the 1920s, the name M. rossiaemeridionalis was forgotten about in the following decades. The discovery in the late 1960s that a population originally assumed to be part of the Common vole M. arvalis actually merited distinction then led to the naming of the new species M. subarvalis Meyer et al. 1972, and it was this population that then proved to be the same thing as M. rossiaemeridionalis.
The third species, the Tatra pine vole Microtus tatricus Kratochvíl, 1952, was first described from Slovakia but is now known to occur in Poland, Rumania and Ukraine. Finally, the Balkan pine vole Microtus felteni Malec & Storch, 1963 is of special interest with regard to recently named European mammals in that it is endemic to the former Yugoslavian province of Macedonia, an area where there are a further two endemic mammals: the Balkan or Stankovic’s mole T. stankovici and the Balkan short-tailed mouse Mus macedonicus. Of the two, the former was only named in 1931 and the latter in 1983, so Macedonia has proved a ‘hot-spot’ for new European mammals.
Another ‘100 years’ vole is the highly distinctive Balkan snow vole or Martino’s snow vole Dinaromys bogdanovi (Martino, 1922), originally named as a species of Microtus but awarded its own genus in 1955. Occurring in Croatia, Bosnia and Herzegovina, it may also be present in Albania and Greece. Fossils show that it formerly occurred more widely in Europe. Finally among voles, there is the Southern water vole Arvicola sapidus Miller, 1908, an endangered species of France, Spain and Portugal.
 Finally among rodents, we come to another obscure and poorly known species, Roach’s mouse-tailed dormouse Myomimus roachi (Bate, 1937). First described from Israel as a fossil, it was discovered in living form in Bulgaria in 1960 and in Turkey in 1991. Several other species of this genus are known, all from eastern Asia, all named during the 20th century [the adjacent image shows one of these, M. personatus of Turkmenistan, Uzbekistan and Iran].
Finally among rodents, we come to another obscure and poorly known species, Roach’s mouse-tailed dormouse Myomimus roachi (Bate, 1937). First described from Israel as a fossil, it was discovered in living form in Bulgaria in 1960 and in Turkey in 1991. Several other species of this genus are known, all from eastern Asia, all named during the 20th century [the adjacent image shows one of these, M. personatus of Turkmenistan, Uzbekistan and Iran].
Moving now to lipotyphlans, or insectivorans or whatever you want to call them, we find that several species have been named within the last 100 years. Europe’s shrew species belong to three genera, Sorex (the long-tailed or red-toothed shrews), Crocidura (the white-toothed shrews) and Neomys (the Old World water shrews), and what’s interesting it that the ‘100 year’ species belong to all three of these. The new Sorex species are the Spanish or Iberian shrew S. granarius Miller, 1910, the Taiga or Even-toothed shrew S. isodon Turov, 1924 and the Appenine shrew S. samniticus Altobello, 1926. Though first described as a subspecies of the Common shrew S. araneus, the Spanish shrew is strongly distinct genetically and in having a particularly unusual short skull. The Taiga shrew occurs from Norway to as far east as Siberia and Sakhalin Island; it is a large, drab species with a broad braincase and particularly narrow snout. Though named in the 1920s it was later regarded as a subspecies of the Dusky shrew S. sinalis, a Chinese species, until Hoffmann (1987) showed that it should have remained as a species. The Appenine shrew is endemic to Italy, and while formerly regarded by some as conspecific with the Common shrew, it is quite different, having a much shorter tail for example.
Moving now to white-toothed shrews, we find that four European species have been named since the 1950s. Shrews have proved very good at colonizing islands, and only within recent decades have mammalogists started to properly describe and differentiate the island endemic white-toothed shrews of the European islands. Crete has its own recently-named white-toothed shrew, the Cretan white-toothed shrew C. zimmermanni Wettstein, 1953, while Pantelleria Island off Italy is home to C. cossyrensis Contoli, 1989. The Pantelleria shrew is controversial, with various studies indicating that it is a subspecies of the Greater white-toothed shrew (C. russula).
During the 1980s two new white-toothed shrews were named from the Canary Islands: the Canary shrew C. canariensis Hutterer et al., 1987 of Fuerteventura, Lanzarote and Lobos, and the Osorio shrew Crocidura osorio Molina & Hutterer, 1989 of Gran Canaria. The only other extant endemic mammal of the Canary Islands, the bat Plecotus teneriffae, was named in 1907 as a subspecies and given species status in 1985, so the islands have proved an important place for the discovery of new European mammals. Incidentally, there were other endemic mammals on the Canary Islands until recently, but they are today extinct. While its discovery falls outside of the last 100 years, of interest is that Sicily’s endemic white-toothed shrew was only named in 1900: the Sicilian shrew C. sicula Miller, 1900.
 Though it has since been demoted to subspecific status, it’s also worth noting that the white-toothed shrew of the Isles of Scilly, Crocidura suaveolens cassiteridum, was originally named as a distinct species (C. cassiteridum) in 1924 (Hinton 1924) [see adjacent image]. This shrew isn’t unique to the Isles of Scilly, as it also occurs on Jersey and Sark, and given that it belongs to a species otherwise restricted to southern Europe it is usually thought of as an introduction from the Mediterranean region. Presumably it made the crossing in fodder or bedding for domestic animals. Incidentally, the Hinton who named the Scilly shrew is Martin Alister Campbell Hinton (1883-1961), former Keeper of Zoology at London’s Natural History Museum, and perhaps best known nowadays as possible perpetrator of the Piltdown hoax.
Though it has since been demoted to subspecific status, it’s also worth noting that the white-toothed shrew of the Isles of Scilly, Crocidura suaveolens cassiteridum, was originally named as a distinct species (C. cassiteridum) in 1924 (Hinton 1924) [see adjacent image]. This shrew isn’t unique to the Isles of Scilly, as it also occurs on Jersey and Sark, and given that it belongs to a species otherwise restricted to southern Europe it is usually thought of as an introduction from the Mediterranean region. Presumably it made the crossing in fodder or bedding for domestic animals. Incidentally, the Hinton who named the Scilly shrew is Martin Alister Campbell Hinton (1883-1961), former Keeper of Zoology at London’s Natural History Museum, and perhaps best known nowadays as possible perpetrator of the Piltdown hoax.
Finally among shrews, there is the Neomys species Miller’s water shrew Neomys anomalus Cabrera, 1907, also known as the Mediterranean or Cabrera or Southern water shrew. In contrast to the better-known Neomys species, N. anomalus is less well adapted for life in water, with a less well-developed tail keel and fewer fringes on the borders of its hind feet, and it differs in the shape of its lower jaw, in penis morphology, and in other characters from the other European Neomys species.
Among ‘100 year’ European lipotyphlans, it’s not all just shrews. Three new European mole species have been named since 1906: the Levant mole Talpa levantis Thomas, 1906, the Iberian mole T. occidentalis Cabrera, 1907, and the Balkan or Stankovic’s mole T. stankovici Martino & Martino, 1931. A fourth species, the Roman mole T. romana Thomas, 1902 was named 104 years ago. While all of these taxa were originally named as distinct species, they later became sunk into the synonymy of other species (yet more examples of laissez-faire lumping: see The many babirusa species: laissez-faire lumping under fire again), only to be resurrected during the 1990s. The Levant mole, an animal known from Bulgaria, Greece, Turkey and the adjacent part of the Caucasus, was mostly regarded as a subspecies of the Mediterranean mole T. caeca, until a revision of 1993, and the Iberian mole was similarly widely regarded as a Mediterranean mole subspecies until 1993. Similarly, the Balkan mole was regarded during recent decades as a subspecies of the Roman mole T. romana.
 Finally, we come to bats. While most European bat species were formally named in the 1800s and before, new taxa continue to be discovered, with several species named this century. Many people might immediately think of the two pipistrelle species dubbed informally the 45 and 55 kHz pipistrelles: in 1993 it was discovered that the ‘species’ Pipistrellus pipistrellus actually consisted of two distinct species, both of which differed in the echolocation frequencies of their calls, and which were later shown to differ in genetics, morphology and behaviour (Barlow et al. 1997, Davidson-Watts & Jones 2006). However, while the many differences between these two species have only recently been acknowledged, both were originally named during the 1700s and 1800s: the 45 kHz pipistrelle is P. pipistellus (Schreber, 1774) while the 55 kHz pipistrelle is P. pygmaeus Leach, 1825. Consequently, neither bat can be considered a ‘100 year’ discovery.
Finally, we come to bats. While most European bat species were formally named in the 1800s and before, new taxa continue to be discovered, with several species named this century. Many people might immediately think of the two pipistrelle species dubbed informally the 45 and 55 kHz pipistrelles: in 1993 it was discovered that the ‘species’ Pipistrellus pipistrellus actually consisted of two distinct species, both of which differed in the echolocation frequencies of their calls, and which were later shown to differ in genetics, morphology and behaviour (Barlow et al. 1997, Davidson-Watts & Jones 2006). However, while the many differences between these two species have only recently been acknowledged, both were originally named during the 1700s and 1800s: the 45 kHz pipistrelle is P. pipistellus (Schreber, 1774) while the 55 kHz pipistrelle is P. pygmaeus Leach, 1825. Consequently, neither bat can be considered a ‘100 year’ discovery.
However, vesper bats have yielded several bona fide new European species within the last 100 years, though as we shall see a few of them are of controversial status. The most recently named of them are the two long-eared bats Plecotus microdontus Spitzenberger et al. 2002 from Austria and P. sardus Mucedda et al., 2002 from Sardinia, though P. microdontus has since been regarded by some as synonymous with the Brown long-eared bat P. auritus. Also recently named is the Alpine long-eared bat P. alpinus Kiefer & Veith, 2001, named for a specimen collected in France in 2001 (Kiefer & Veith 2001). Additional specimens are known from Greece, Liechtenstein, Austria, Croatia and Switzerland, so there is every indication that the species is widespread. The Croatian specimen was collected in 1972 and the specimen from Liechtenstein in 1961: a reminder that the actual ‘discovery’ date of a species often doesn’t match the time when it becomes technically named and/or described.
 Yet another recently recognised species, P. macrobullaris Kuzjakin, 1965, was named for long-eared bats from Switzerland and Austria supposedly intermediate between the Brown long-eared bat and Grey long-eared bat P. austriacus, but shown by Spitzenberger et al. (2001) to be worthy of species status. P. macrobullaris is now known from Croatia and elsewhere. To confuse matters further, recent work (see Juste et al. 2004) indicates that both P. microdontus and P. alpinus are synonymous with P. macrobullaris [adjacent image shows a long-eared bat. And no, I have no idea what species it is].
Yet another recently recognised species, P. macrobullaris Kuzjakin, 1965, was named for long-eared bats from Switzerland and Austria supposedly intermediate between the Brown long-eared bat and Grey long-eared bat P. austriacus, but shown by Spitzenberger et al. (2001) to be worthy of species status. P. macrobullaris is now known from Croatia and elsewhere. To confuse matters further, recent work (see Juste et al. 2004) indicates that both P. microdontus and P. alpinus are synonymous with P. macrobullaris [adjacent image shows a long-eared bat. And no, I have no idea what species it is].
Several new long-eared bat subspecies have also been named within the last few decades, and new data has caused some of them to be newly elevated to species level. Within P. auritus, the subspecies P. a. hispanicus (later reidentified as a subspecies of P. austriacus) was named in 1957, P. a. kolombatovici in 1980, and P. a. begognae in 1990. Genetic studies have shown that P. a. kolombatovici is distinct enough to be regarded as a full species (Mayer & von Helverson 2001, Spitzenberger et al. 2001), though the animal labelled as P. a. kolombatovici by Spitzenberger et al. (2001) later turned out to be P. alpinus. Another form first named as a subspecies of P. auritus, P. a. teneriffae Barret-Hamilton, 1907, was recognised as worthy of species status in 1985. Though they started their taxonomic histories as subspecies, both P. kolombatovici and P. teneriffae can therefore be stated to have been discovered within the last 100 years.
Another new vesper bat, this time a mouse-eared bat, is in the ‘100 years’ club, but it seems unlikely to be a valid species. It’s the Nathaline bat Myotis nathalinae Tupinier, 1977, described for two specimens from Ciudad Real in Spain. However, it’s highly similar genetically and morphologically to Daubenton’s bat M. daubentonii (Tupinier 1977). Indeed Bogdanowicz (1990) found that the skull morphology of M. nathalinae fell within the range of variation exhibited by M. daubentonii populations, and therefore argued against the idea that it should be regarded as a valid species, while genetic samples of M. nathalinae have also fallen within the range of variation exhibited by M. daubentonii (Mayer & von Helverson 2001). Other studies have produced the same result, so bat workers generally regard M. nathalinae as a subspecies of M. daubentonii.
A second new mouse-eared bat, M. alcathoe von Helverson et al., 2001, is morphologically and genetically distinct, and noteworthy in being Europe’s smallest mouse-eared bat, and the one with the most high-pitched echolocation calls. First reported from Greece and Hungary, in 2003 it was reported from Slovakia.
So, so far it’s all been rodents, insectivores and bats: exactly those groups of mammals you’d expect to contain recently-discovered species. Indeed, that is about it. There is, however, a ‘100 year’ European lagomorph: the Broom hare Lepus castroviejoi Palacios, 1977 of the Catabrian Mountains of north-west Spain, a species regarded as merely a population of the European hare L. europaeus until 1976 (Palacios 1977). This poorly known hare is obscure and has been widely overlooked, in fact it’s missing from several (post-1977!) field guides on European mammals. There does appear to be widespread acceptance of its specific status, however, even though there is some indication that the species hybridizes with the Mountain hare L. timidus (Melo-Ferreira et al. 2005).
It’s pretty clear then that the Cypriot mouse is most certainly not ‘the first new mammal species to be found in Europe in over a century’, and I’m amazed that such a claim has been made. Despite the message that journalists write into their stories all the time, the discovery of new species is a routine thing, not an extraordinary one, and that goes even for mammals, and even for Europe. Don’t get me wrong: the Cypriot mouse is still a very interesting and significant discovery, but it is clearly not the major scientific event that has been implied by some.
To conclude, those European mammals named within the past 100 years – excluding the Cypriot mouse – are as follows. I might have missed some, in which case please let me know [UPDATE: list ammended as of 18-11-2006. Thanks to those who have provided new data]. As noted, a few species are of dubious status, and have been marked with **.
- Cretan spiny mouse Acomys minous Bate, 1906
- Western house mouse Mus domesticus Schwartz & Schwartz, 1943
- Alpine wood mouse Apodemus alpicola Heinrich, 1952
- Mount Hermon field mouse A. iconicus Heptner, 1948
- Balkan short-tailed mouse Mus macedonicus Petrov & Ruzic, 1983
- Bavarian pine vole Microtus bavaricus Konig, 1962
- Cabrera’s vole M. cabrerae Thomas, 1906
- Sibling vole M. rossiaemeridionalis Ognev, 1924
- Tatra pine vole M. tatricus Kratochvíl, 1952
- Balkan pine vole M. felteni Malec & Storch, 1963
- Balkan snow vole or Martino’s snow vole Dinaromys bogdanovi (Martino, 1922)
- Southern water vole Arvicola sapidus Miller, 1908
- Roach’s mouse-tailed dormouse Myomimus roachi (Bate, 1937)
- Spanish or Iberian shrew Sorex granarius Miller, 1910
- Taiga or Even-toothed shrew S. isodon Turov, 1924
- Appenine shrew S. samniticus Altobello, 1926
- Cretan white-toothed shrew Crocidura zimmermanni Wettstein, 1953
- Pantelleria Island shrew C. cossyrensis Contoli, 1989 **
- Canary shrew C. canariensis Hutterer et al., 1987
- Osorio shrew C. osorio Molina & Hutterer, 1989
- Miller’s water shrew Neomys anomalus Cabrera, 1907
- Levant mole Talpa levantis Thomas, 1906
- Iberian mole T. occidentalis Cabrera, 1907
- Balkan or Stankovic’s mole T. stankovici Martino & Martino, 1931
- Alpine long-eared bat Plecotus alpinus Kiefer & Veith, 2001 **
- P. microdontus Spitzenberger et al. 2002 **
- P. kolombatovici (Dulic, 1980)
- P. teneriffae Barret-Hamilton, 1907
- P. macrobullaris Kuzjakin, 1965
- Nathaline bat Myotis nathalinae Tupinier, 1977 **
- M. alcathoe von Helverson et al., 2001
- Broom hare Lepus castroviejoi Palacios, 1977
For the latest news on Tetrapod Zoology do go
here.
Refs - -
Barlow, K. E., Jones, G. & Barratt, E. M. 1997. Can skull morphology be used to predict ecological relationships between bat species? A test using two cryptic species of pipistrelle. Proceedings of the Royal Society of London B 264, 1695-1700.
Bogdanowicz, W. 1990. Geographic variation and taxonomy of Daubenton’s bat, Myotis daubentoni, in Europe. Journal of Mammalogy 71, 205-218.
Davidson-Watts, I. & Jones, G. 2005. Differences in foraging behaviour between Pipistrellus pipistrellus (Schreber, 1774) and Pipistrellus pygmaeus (Leach, 1825). Journal of Zoology 268, 55-62.
Hinton, M. A. C. 1924. On a new species of Crocidura from Scilly. Annals and Magazine of Natural History 14, 509-510.
Hoffmann, R. S. 1987. A review of the systematics and distribution of Chinese red-toothed shrews (Mammalia: Soricinae). Acta Theriologica Sinica 7, 100-139.
Juste, J., Ibáñez, C., Muñoz, J., Trujillo, D., Benda, P., Karatş, A. & Ruedi, M. 2004. Mitochondrial phylogeography of the long-eared bats (Plecotus) in the Mediterranean Palaearctic and Atlantic Islands. Molecular Phylogenetics and Evolution 31, 1114-1126.
Kiefer, A. & Veith, M. 2001. A new species of long-eared bat from Europe (Chiroptera: Vespertilionidae). Myotis 39, 5-16.
Kryštufek, B. & Mozetič Francky, B. 2005. Mt. Hermon field mouse Apodemus iconicus is a member of the European mammal fauna. Folia Zoologica 54, 69-74.
Mayer, F. & von Helversen, O. 2001. Cryptic diversity in European bats. Proceedings of the Royal Society of London B 268, 1825-1832.
Melo-Ferreira, J., Boursot, P., Suchentrunk, F., Ferrand, N. & Alves, P. C. 2005. Invasion from the cold past: extensive introgression of mountain hare (Lepus timidus) mitochondrial DNA into three other hare species in northern Iberia. Molecular Ecology 14, 24-59.
Palacios, F. 1977. Descripcion de una nueva especie de liebre (Lepus castroviejoi) endémica de la cordillera Cantabrica. Doñana Acta Vertebrata 3, 205-223.
Spitzenberger, F., Piálek, J. & Haring, E. 2001. Systematics of the genus Plecotus (Mammalia, Vespertilionidae) in Austria based on morphometric and molecular investigations. Folia Zoologica 50, 161-172.
Tupinier, Y. 1977. Description d'une Chauve-souris nouvelle: Myotis nathalinae nov. sp. (Chiroptera, Vespertilionidae). Mammalia 41, 327-340.

 I’ve always been very interested in phorusrhacids and, unlike many of the animals I write about (the shame), I have some experience with them. What are they? They are universally agreed to be relatives of the living seriemas (Cariamidae), but differ from them in having a far more robust bill and jaws, smaller bony processes on the humerus, and a narrower pelvis. They also, of course, grew to a much larger size. The two living seriema species are South American, but members of similar, closely related groups (the bathornithids and idiornithids) inhabited
I’ve always been very interested in phorusrhacids and, unlike many of the animals I write about (the shame), I have some experience with them. What are they? They are universally agreed to be relatives of the living seriemas (Cariamidae), but differ from them in having a far more robust bill and jaws, smaller bony processes on the humerus, and a narrower pelvis. They also, of course, grew to a much larger size. The two living seriema species are South American, but members of similar, closely related groups (the bathornithids and idiornithids) inhabited 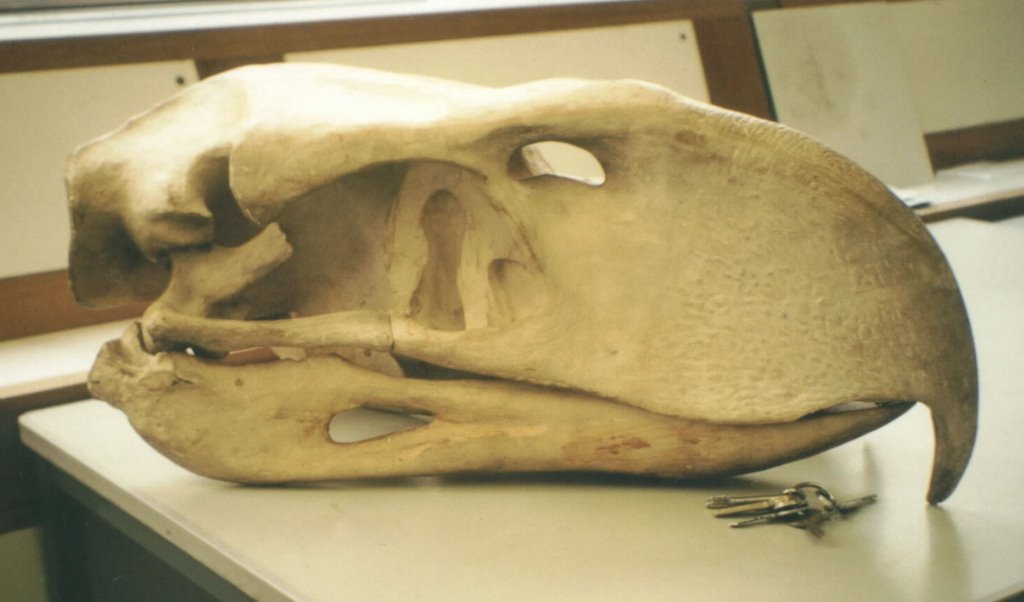 It is of minor frustration that the phorusrhacids we hear about the most are among the most poorly known. The ‘best known’ phorusrhacid, the one featured in every single prehistoric animal book, is Phorusrhacos longissimus from the Miocene of Argentina. But it’s only ‘best known’ because it was the first member of the group to be named, and compared to a number of far more obscure species, it is poorly known and mysterious. Of its skull, for example, we only have the lower jaw and some fragments of cranium. Florentino Ameghino (1854-1911), the famous Argentine zoologist/palaeontologist who discovered and named it and several other phorusrhacids, did write in 1895 of seeing a complete skull, encased in rock in the field, but he was only able to sketch it and recover fragments. His drawing is of a complete, pristine skull and it is on the basis of this that an entire replica skull has been produced (see accompanying image). Compare this with the patagornithines Patagornis and Andalgalornis, for example, both of which are known from awesome, complete skulls with good, associated, near-complete skeletons.
It is of minor frustration that the phorusrhacids we hear about the most are among the most poorly known. The ‘best known’ phorusrhacid, the one featured in every single prehistoric animal book, is Phorusrhacos longissimus from the Miocene of Argentina. But it’s only ‘best known’ because it was the first member of the group to be named, and compared to a number of far more obscure species, it is poorly known and mysterious. Of its skull, for example, we only have the lower jaw and some fragments of cranium. Florentino Ameghino (1854-1911), the famous Argentine zoologist/palaeontologist who discovered and named it and several other phorusrhacids, did write in 1895 of seeing a complete skull, encased in rock in the field, but he was only able to sketch it and recover fragments. His drawing is of a complete, pristine skull and it is on the basis of this that an entire replica skull has been produced (see accompanying image). Compare this with the patagornithines Patagornis and Andalgalornis, for example, both of which are known from awesome, complete skulls with good, associated, near-complete skeletons.  The new phorusrhacid described by Chiappe & Bertelli (2006) consists only of a skull and some leg bones (other elements might be known, but aren’t mentioned), but is significant for its size and the completeness of the skull. Discovered in Miocene rocks of
The new phorusrhacid described by Chiappe & Bertelli (2006) consists only of a skull and some leg bones (other elements might be known, but aren’t mentioned), but is significant for its size and the completeness of the skull. Discovered in Miocene rocks of